
Page 18 - Fall2012
P. 18
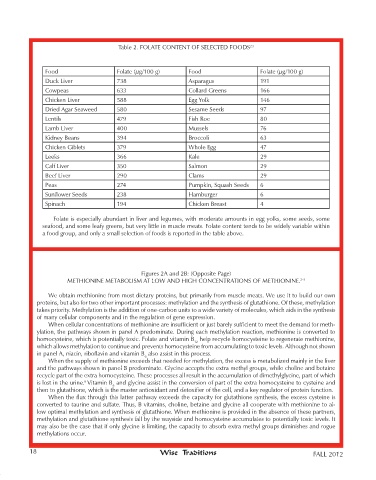
Table 2. FOLATE CONTENT OF SELECTED FOODS 23
Food Folate (µg/100 g) Food Folate (µg/100 g)
Duck Liver 738 Asparagus 191
Cowpeas 633 Collard Greens 166
Chicken Liver 588 Egg Yolk 146
Dried Agar Seaweed 580 Sesame Seeds 97
Lentils 479 Fish Roe 80
Lamb Liver 400 Mussels 76
Kidney Beans 394 Broccoli 63
Chicken Giblets 379 Whole Egg 47
Leeks 366 Kale 29
Calf Liver 350 Salmon 29
Beef Liver 290 Clams 29
Peas 274 Pumpkin, Squash Seeds 6
Sunflower Seeds 238 Hamburger 6
Spinach 194 Chicken Breast 4
Folate is especially abundant in liver and legumes, with moderate amounts in egg yolks, some seeds, some
seafood, and some leafy greens, but very little in muscle meats. Folate content tends to be widely variable within
a food group, and only a small selection of foods is reported in the table above.
Figures 2A and 2B: (Opposite Page)
METHIONINE METABOLISM AT LOW AND HIGH CONCENTRATIONS OF METHIONINE.
1-3
We obtain methionine from most dietary proteins, but primarily from muscle meats. We use it to build our own
proteins, but also for two other important processes: methylation and the synthesis of glutathione. Of these, methylation
takes priority. Methylation is the addition of one-carbon units to a wide variety of molecules, which aids in the synthesis
of many cellular components and in the regulation of gene expression.
When cellular concentrations of methionine are insufficient or just barely sufficient to meet the demand for meth-
ylation, the pathways shown in panel A predominate. During each methylation reaction, methionine is converted to
homocysteine, which is potentially toxic. Folate and vitamin B help recycle homocysteine to regenerate methionine,
12
which allows methylation to continue and prevents homocysteine from accumulating to toxic levels. Although not shown
in panel A, niacin, riboflavin and vitamin B also assist in this process.
6
When the supply of methionine exceeds that needed for methylation, the excess is metabolized mainly in the liver
and the pathways shown in panel B predominate. Glycine accepts the extra methyl groups, while choline and betaine
recycle part of the extra homocysteine. These processes all result in the accumulation of dimethylglycine, part of which
is lost in the urine. Vitamin B and glycine assist in the conversion of part of the extra homocysteine to cysteine and
6
6
then to glutathione, which is the master antioxidant and detoxifier of the cell, and a key regulator of protein function.
When the flux through this latter pathway exceeds the capacity for glutathione synthesis, the excess cysteine is
converted to taurine and sulfate. Thus, B vitamins, choline, betaine and glycine all cooperate with methionine to al-
low optimal methylation and synthesis of glutathione. When methionine is provided in the absence of these partners,
methylation and glutathione synthesis fall by the wayside and homocysteine accumulates to potentially toxic levels. It
may also be the case that if only glycine is limiting, the capacity to absorb extra methyl groups diminishes and rogue
methylations occur.
18 Wise Traditions FALL 2012 FALL 2012 Wise Traditions
101665_text.indd 18 9/14/12 1:33 AM